翻译
在遗传信息的传递过程中,DNA将其遗传信息转录给mRNA,这是基因表达的第一步,mRNA再指导蛋白质的合成,这是基因表达的第二步,即翻译(translation)。
翻译过程需要300多种生物大分子的参与,其中包括核糖体、mRNA、tRNA及多种蛋白质因子。mRNA作为翻译的模板,决定蛋白质中的氨基酸顺序;tRNA作为运输工具,携带氨基酸准确进入指定位置;rRNA与多种蛋白质组成核糖体,作为蛋白质合成的装配机器。
一、翻译的基本原理
(一)mRNA
生物体的遗传信息储存于DNA分子四种碱基的排列顺序中,通过转录,这种顺序转移到RNA分子中。RNA链上从5'端到3'端每3个相邻的核苷酸可以决定一个特定的氨基酸,这种核苷酸三联体被称为密码子(codon)。除了5'端和3'端的非翻译区之外,整个mRNA链即是由串联排列的密码子组成,这样,就把mRNA上的碱基排列顺序叫做遗传密码(genetic code)。在蛋白质合成中,它们决定氨基酸的排列顺序。核苷酸有4种,每3个为一组,共可组成64种密码子(表9-2)。
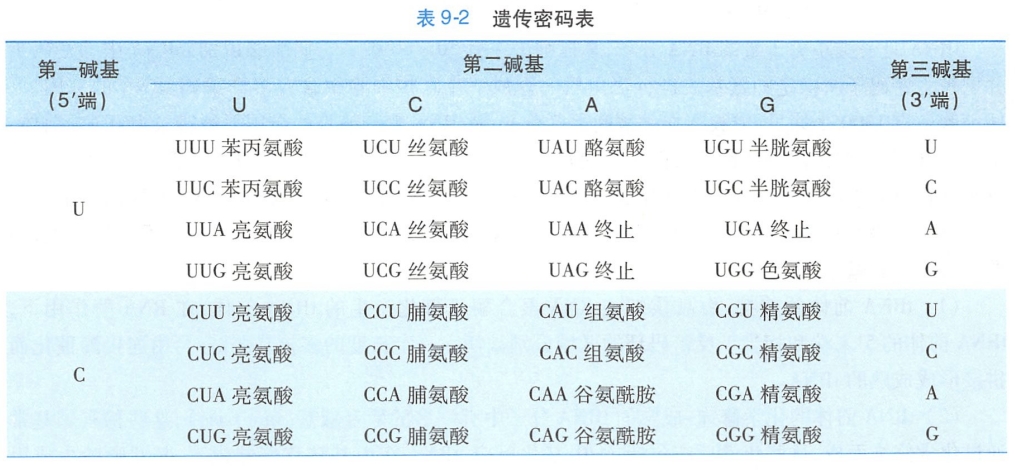
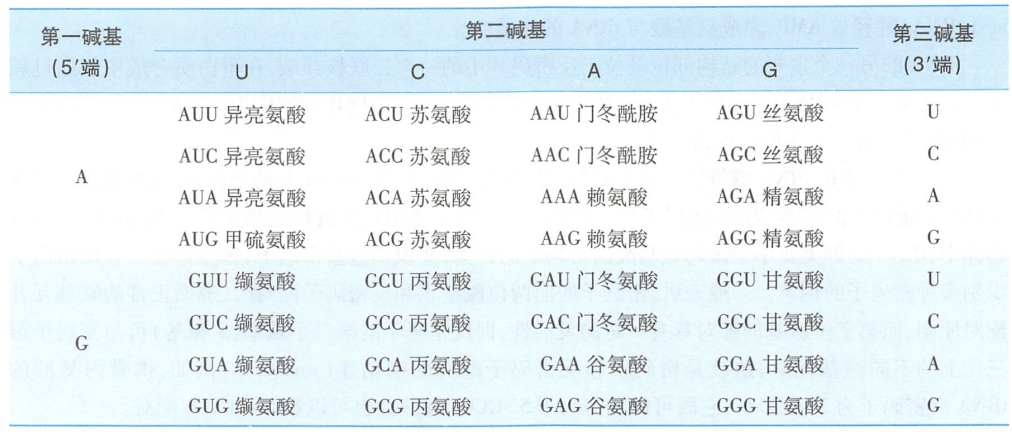
遗传密码有如下特点:
- 通用性(universal):从原核生物到真核生物,几乎所有生物体中的遗传密码都是通用的,就是说,一个特定的碱基序列无论在哪一种生物体中均编码同一种氨基酸。但也有例外,动物细胞线粒体和植物细胞叶绿体中,蛋白质合成所使用的三联密码子有少数与目前通用密码子有所不同(详见第六章)。
- 简并性(degeneracy):密码子有64种,其中三个密码子UAA、UAG、UGA为终止密码,不决定氨基酸,其余61个密码子都可以编码氨基酸,而氨基酸只有20种,所以,必然出现多个密码子决定同一氨基酸的情况,这种现象叫做遗传密码的简并性。
- 连续性(commaless):在翻译过程中,遗传密码的阅读是连续的,在mRNA上的每个三联体密码子之间没有间隔。
- 方向性(direction): mRNA中每个密码子的阅读方向必须从5'→3',因而起始密码子AUG总是位于5'端,终止密码子则位于3'端,翻译过程是从mRNA的5'→3'方向进行。
(二)tRNA
tRNA既能够识别mRNA上的密码子,又能携带特定的氨基酸,因而被称为蛋白质合成的接合器(adaptor)(图9-9)。
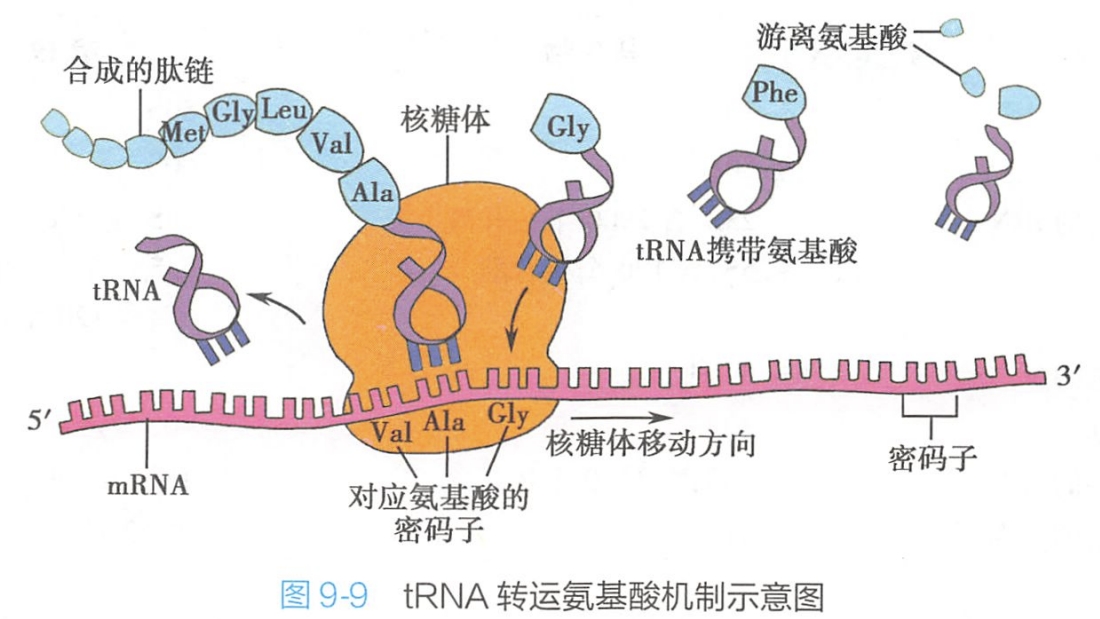
tRNA结构的主要特点是3'端的CCA序列,活化后的氨基酸就是通过CCA序列上的-OH与tRNA结合,然后被带到核糖体上参与蛋白质的合成。
1.携带特定氨基酸
tRNA携带特定氨基酸的过程是通过氨酰-tRNA合成酶催化的两步反应完成的: #生化
- 反应的第一步是氨基酸与ATP在氨酰-RNA合成酶作用下形成氨酰-AMP,完成氨基酸的活化;
- 第二步,活化氨基酸的氨酰基被转移至tRNA分子上形成氨酰-tRNA(amino-acyl-tRNA)并释放AMP,完成氨基酸与RNA的连接。
2.反密码子
tRNA的另一个重要的结构部位是位于反密码环中的一个三联核苷酸,在蛋白质合成中能通过碱基互补配对识别mRNA上的密码子,这种存在于tRNA中的三联核苷酸被称为反密码子(anticodon)。
不同的tRNA分子有不同的反密码子。在翻译过程中,tRNA携带特定的氨基酸,通过反密码子识别mRNA上的密码子,实现遗传信息从mRNA到蛋白质的传递。
3.摆动性
在生物体中,tRNA的种类要小于编码氨基酸的密码子的种类,要识别这些密码子,必然存在一种反密码子识别多种密码子的现象。一般来讲,密码子的前两位碱基在和反密码子配对时,遵循正常的碱基互补配对原则,而第三位碱基的配对具有一定的灵活性,即反密码子的第三个碱基(5'碱基)可与密码子第三位上的不同碱基配对,这就是密码子和反密码子配对的摆动性(wobble)。例如,携带丙氨酸的tRNA反密码子为3'-CGC-5',它既可以和密码子5'-GCG-3'配对,也可以和5'-GCU-3'配对。
二、核糖体
核糖体(ribosome)也称核蛋白体(与核糖核蛋白颗粒应该有区别,核仁的颗粒组分)。核糖体几乎存在于所有的细胞内,即使是最简单的支原体细胞也至少含有上百个核糖体。此外,在线粒体中也含有核糖体。
(一)核糖体组成
原核细胞是70S的核糖体,真核细胞是80S的核糖体。真核细胞线粒体内的核糖体近于70S。
70S和80S的核糖体均由大小不同的两个亚单位组成,大的称为大亚基,小的称为小亚基。核糖体大小亚基在细胞内一般以游离状态存在,只有当小亚基与RNA结合后,大亚基才与小亚基结合,形成完整的核糖体。
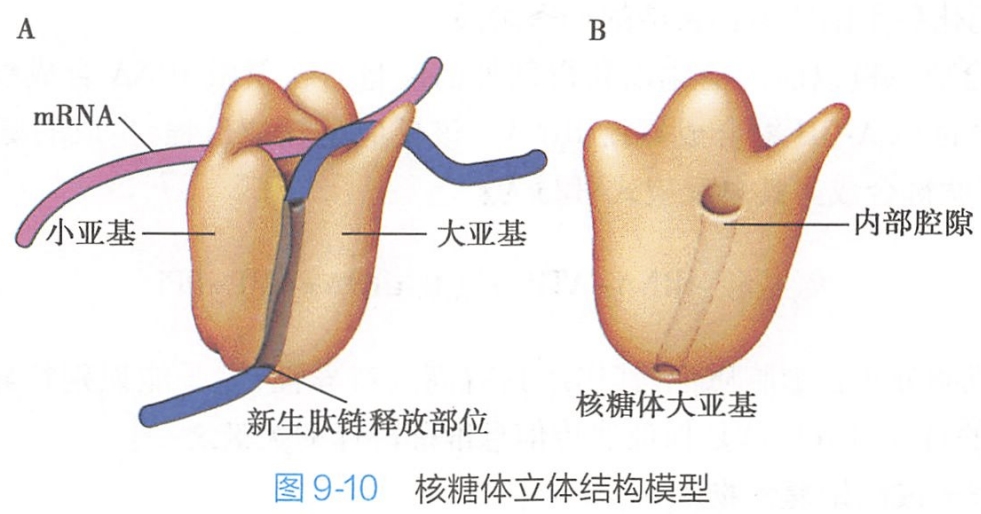
在70S核糖体中:
小亚基30S=16S rRNA+21种蛋白质
大亚基50S=23S rRNA+5S rRNA+34种蛋白质
在80S核糖体中:
小亚基40S=18S rRNA+33种蛋白质
大亚基60S=28S rRNA+5.8S rRNA+5S rRNA+49种蛋白质(表9-3)。

RNA约占核糖体的60%,蛋白质约占40%。核糖体中的RNA主要构成核糖体的骨架,将蛋白质串联起来,并决定蛋白质的定位。
(二)核糖体功能
原核细胞:除少数核糖体附着在质膜以外,大部分核糖体则以游离形式存在。
真核细胞:很多核糖体附着在内质网膜的外表面,形成糙面内质网,还有一部分核糖体以游离形式分布在细胞质溶胶内。
呈游离状态的核糖体称为游离核糖体,附着在膜上的核糖体称为附着核糖体。不同点在于所合成的蛋白质种类不同,如游离核糖体主要合成细胞内的某些基础性蛋白,附着核糖体主要合成细胞的分泌蛋白和膜蛋白。
核糖体上存在多个与蛋白质的多肽链形成密切相关的活性部位(图9-11),主要有:
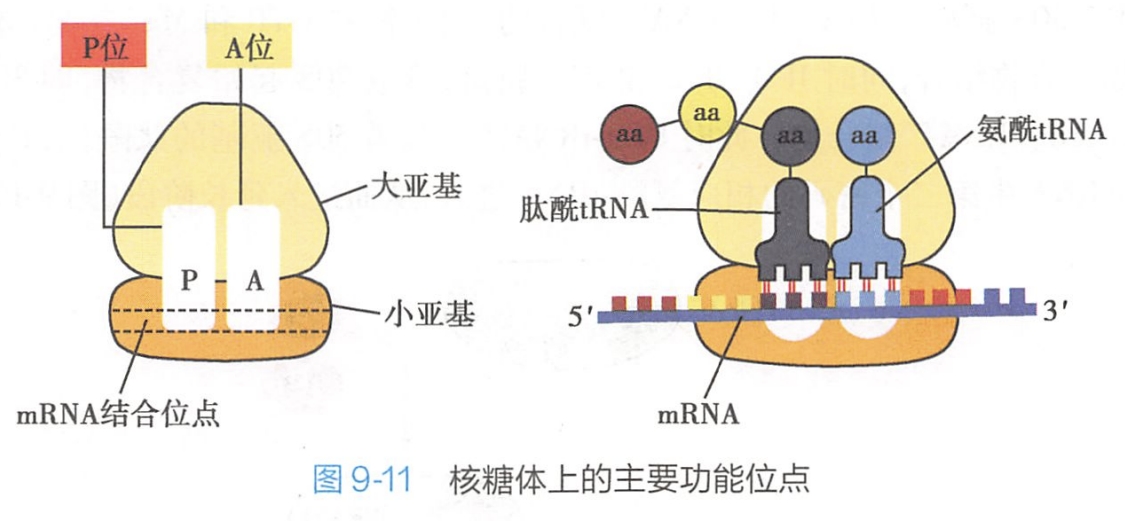
1.核糖体结合位点(原核)
原核生物30S小亚基具有专一性地识别和选择mRNA起始位点的性质。
30S小亚基通过其16S rRNA的3'端与mRNA5'端起始密码子上游碱基配对结合。在原核生物的mRNA5'端起始密码子的上游都有一个SD序列(Shine-Dalgarno sequence),即5'-AGGAGGU-3'序列,这个富嘌呤区与30S小亚基上16S rRNA3'端的富嘧啶区序列5'-CACCUCCUUA-3'互补。SD序列可指导 mRNA的起始密码子正确定位在30S小亚基的P位,SD序列又称为核糖体结合位点(ribosomal binding site,RBS)。
在真核生物中,核糖体上有专一位点或因子识别mRNA的帽子结构,使RNA与核糖体结合。
2.P位(peptidyl site)
是肽酰tRNA结合的位置。它大部分位于小亚基,小部分位于大亚基,是结合起始肽酰-RNA并向A位给出氨基酸的位置。
3.A位(aminoacyl site)
是氨酰tRNA结合的位置。它大部分位于大亚基而小部分位于小亚基,是结合一个新进入的氨酰tRNA的位置。
4.转肽酶(transpeptidase)活性部位
位于P位和A位的连接处,其作用是在肽链延长时,催化进入核糖体的氨基酸之间形成肽键。
5.参与蛋白质合成的因子的结合部位
如结合起始因子(initiation factor,IF)、延长因子(elongation factor,EF)和终止因子或释放因子(release factor,RF)的部位。
三、蛋白质合成过程(原核)
蛋白质生物合成可分为五个阶段,即氨基酸的活化、多肽链合成的起始、肽链的延长、肽链的终止和释放、蛋白质合成后的加工修饰。(你这都说是合成后的加工了。)
(一)氨基酸活化
氨基酸在参与合成肽链以前需活化以获得额外的能量。在氨酰-tRNA合成酶作用下,氨基酸的羧基与tRNA3'末端的CCA-OH缩合成氨酰-tRNA。
氨酰:氨基酸分子去掉羟基后,剩下的一价原子团统称为氨酰。
伯酰胺:-CONH2就是酰胺基。
仲酰胺:羰基连上一个-(NH)-就是酰胺键,-(C=O)-(NH)-。两端都还可以连接烃基R。
叔酰胺:羰基连上一个-(N)--也是酰胺键,-(C=O)-N--。C上连一个烃基,N上可以连两个烃基。
氨基酸与tRNA的连接分为两个步骤。(当然,其实是一种亲核加成)
- 首先是氨基酸和ATP缩合形成氨基酸与腺苷酸的混合酸酐中间体,
- 然后中间体非共价结合至活性位点。
- 第二步是氨酰基转移反应,氨基酸被酯化到tRNA3'末端的A的2'或3'-OH。
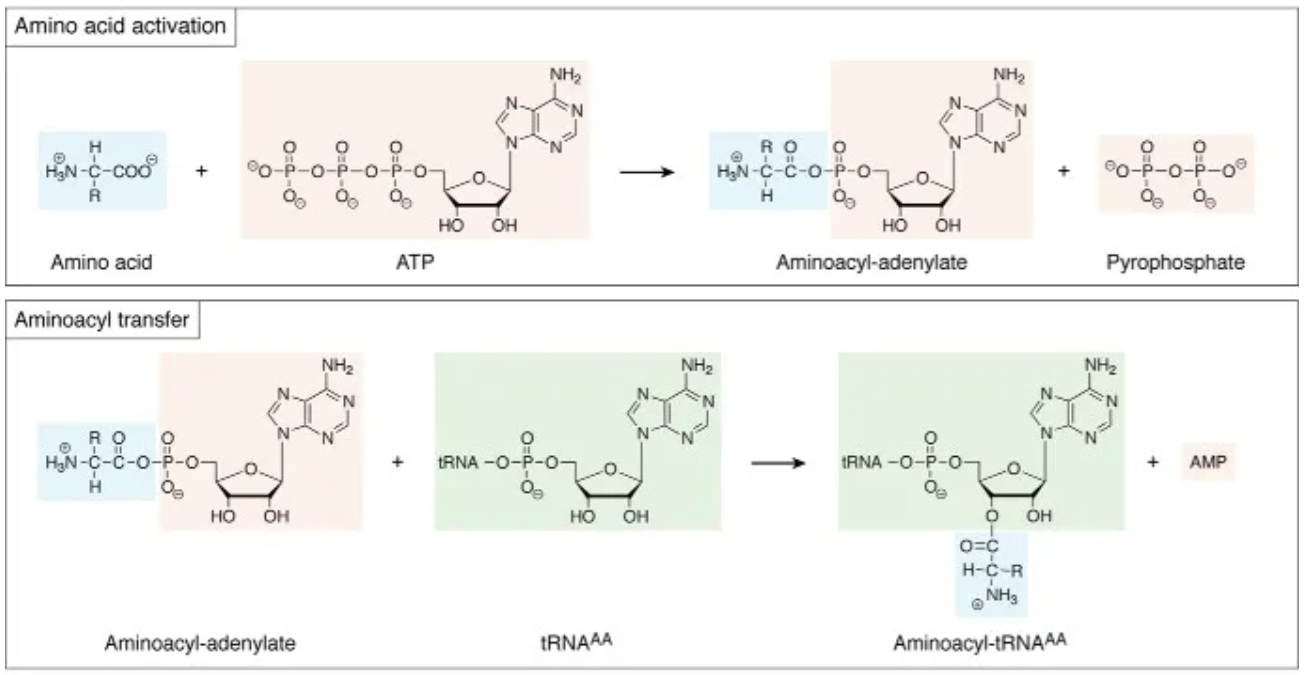
氨酰tRNA合成酶(ARS)分为两类,每类各10种。两类之间在结构和功能上都有差别,例如I类酶在tRNA的2'-OH上氨基酰化,而II类酶通常在3'-OH上氨基酰化。
该反应是耗能过程,生成的氨酰-tRNA中酯酰键含较高能量,可用于肽键合成。其总反应式可写为:氨基酸+tRNA+ATP→氨酰-tRNA+AMP+PPi
氨酰-tRNA合成酶分布在细胞质中,其作用具有高度特异性,它既能识别特异的氨基酸,又能辨认携带该种氨基酸的特异tRNA。
(二)起始过程形成起始复合物
起始阶段指核糖体大、小亚基,mRNA和具有启动作用的起始氨酰-tRNA装配为起始复合物的过程。
具有启动作用的氨酰-tRNA在原核细胞是甲酰甲硫氨酰-tRNA(fMet-tRNA),在真核细胞是甲硫氨酰-tRNA(Met-tRNAM)。
在大肠杆菌中翻译起始复合物形成的过程:
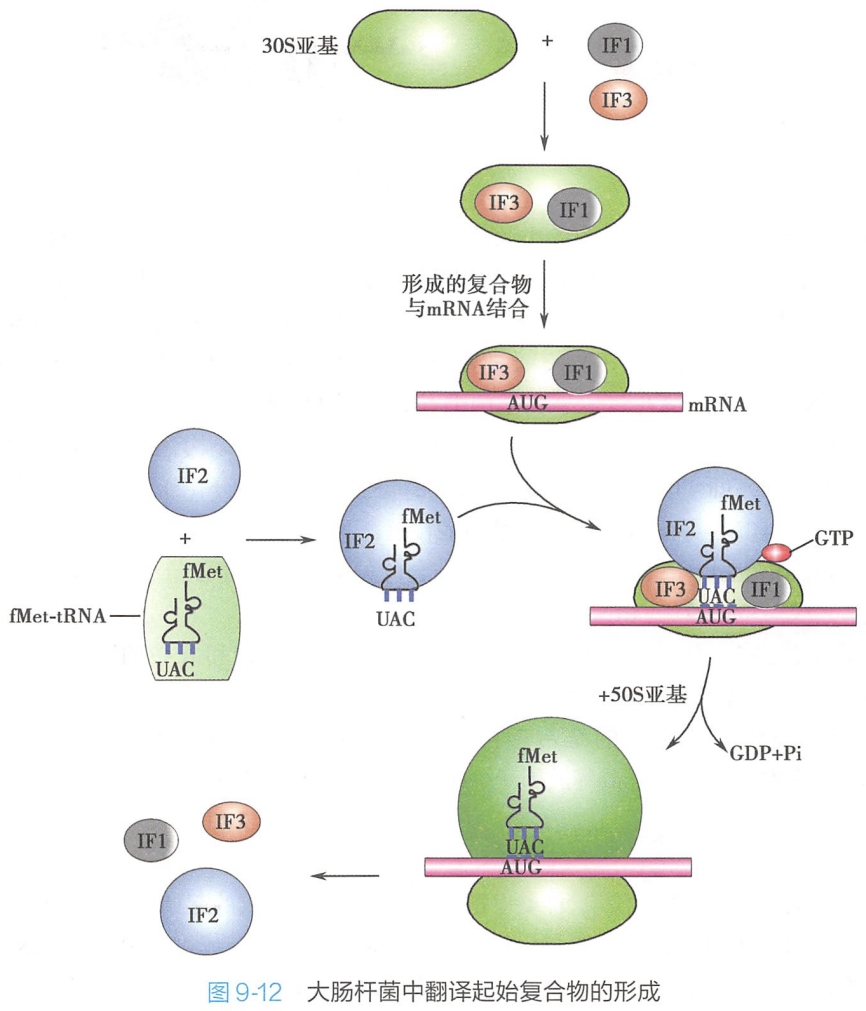
- 在起始因子-1(IF-1)和IF-3作用下,核糖体30S小亚基附着于mRNA起始信号部位(SD序列),形成IF1-IF3-30S亚基-mRNA复合物;
- 在起始因子-2作用下,fMet-tRNA与mRNA分子中的AUG相结合(密码子与反密码子配对),形成30S前起始复合物,即IF1-IF2-IF3-30S亚基-mRNA-fMet-tRNA复合物,此步需要GTP和Mg2+参与;
- 50S亚基与上述的30S前起始复合物结合,同时IF-1、IF-2和IF-3脱落,形成70S起始复合物,即30S亚基-mRNA-50S亚基-mRNA-fMet-tRNAfMet复合物。(哥们你认真的吗?)此时fMet-tRNAfMet占据着肽酰位(P位),而A位留空有待于对应mRNA中第二个密码的相应氨酰-tRNA进入,从而进入延长阶段(图9-12)。
与原核细胞相比,真核细胞蛋白质合成起始过程更为复杂。参与该过程的起始因子被称为eIF(e代表真核细胞)。某些eIF参与了40S小亚基与mRNA5'端帽子结构的结合。在起始复合物形成后,40S小亚基在mRNA上扫描直到发现正确的AUG起始密码子,然后eIF被释放,60S大亚基结合到起始复合物上来,启动蛋白质的合成。此外,mRNA3'端尾部的poly-A可以促进起始复合物的形成,提高翻译效率。
(三)肽链延长是多因子参与的核糖体循环过程
起始复合物形成后,根据mRNA上密码子序列的指导,各种氨酰-tRNA依次结合到核糖体上使肽链从N端向C端逐渐延长。由于肽链延长在核糖体上连续循环进行,所以这个过程又称为核糖体循环(ribosome circula tion)。
核糖体循环包括进位(registration)、成肽(peptide bond formation)、转位(translocation)三个步骤(图9-l3)。每经过一个循环肽链增加一个氨基酸残基。
1.进位
起始复合物形成时,P位已被起始-tRNA占据,而A位是空的。根据70S起始复合物A位上 mRNA密码子,特异的氨酰-tRNA进入A位。
此步骤需 GTP、Mg2+和延长因子EF-Tu与EF-Ts。EF-Tu和EF-Ts是延长因子EF-T的两个亚基。
- EF-Tu与GTP、氨酰-tRNA反应生成三元复合物——氨酰-tRNA-EF-Tu-GTP,
- 该复合物中tRNA的反密码子与小亚基上mRNA结合后,其中的GTP分解释放Pi,EF-Tu-GDP脱落
- 并与EF-Ts反应生成GDP和EF-Tu-EF-Ts,
- 后者再与GTP反应,释放出EF-Ts生成EF-Tu-GTP并进入下一次延长反应。
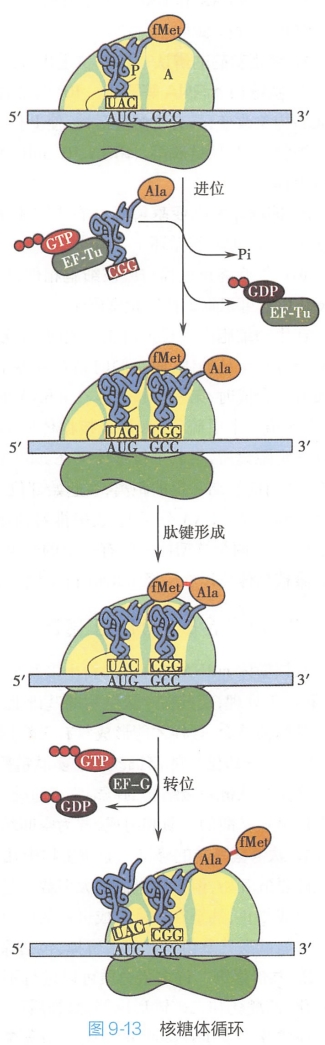
2.成肽
在起始复合物形成后,核糖体的P位上已结合了fMet-tRNA。在进位后,P位和A位上各结合了一个氨酰tRNA。
两个氨基酸之间在核糖体大亚基转肽酶的作用下,P位上的氨基酸提供-COOH基(-C=O),与A位上的氨基酸的-NH2基形成肽键,从而使P位上的氨基酸连接到A位氨基酸的氨基上,这就是成肽。 #?
成肽后,在A位上形成了一个二肽酰RNA。P位上的tRNA随之从核糖体上脱落下来,使P位空出。
3.转位
P位空出后,在延长因子G(EF-G)的作用下,由GTP供能,核糖体沿mRNA5'端向3'端移动一个密码子距离,结果使肽酰tRNA由A位移到P位,空出的A位可接受新的氨酰-RNA。
真核细胞的肽链延长过程与原核细胞大致相同,仅在参与的因子方面有所区别。如真核细胞的延长因子有3种,分别为eEF1α、eEF1βγ和eEF2;转位时所需因子为eEF-2。( #? 我咋有点不信呢)
(四)肽链合成终止过程
包括三个步骤在核糖体向mRNA3'端移动中,肽链也逐渐延长,当mRNA的终止信号UAA、UAG、UGA中任一种进入A位时,没有任何tRNA能与之识别,只有释放因子(RF)识别这种信号。原核细胞的RF有3种,RF-1识别UAA和UAG,RF-2识别UAA和UGA,RF-3结合GTP并促进RF-1、RF-2与核糖体结合。终止的过程如下:
1.终止密码的辨认
当A位上出现终止密码时,RF-1或RF-2识别并结合到A位上。
2.肽链和mRNA等释出
RF的结合使核糖体上转肽酶构象改变,具有水解酶活性,使P位上tRNA与肽链间酯键水解,肽链脱落。tRNA、RF、mRNA也随后从核糖体上释出。
3.核糖体大小亚基解聚
在RF-3作用下,大小亚基解聚,重新进入新循环。
真核细胞终止过程与原核细胞相似,但其释放因子为eRF1,它可识别3种终止密码子。
在生物细胞内进行蛋白质合成时,每分子mRNA上不只结合一个核糖体,而是同时结合着多个核糖体。蛋白质开始合成时,第一个核糖体在mRNA的起始部位结合,引入第一个甲硫氨酸,接着核糖体向RNA的3'端移动一定距离后,第二个核糖体又在mRNA的起始部位结合,并向前移动一定的距离,在起始部位又结合第三个核糖体,依次下去直至终止。这样,多个核糖体结合到一个mRNA分子上,成串排列,形成蛋白质合成的功能单位,称为多聚核糖体(polyribosome)(图9-14)。两个核糖体之间有一定的长度间隔,每个核糖体都独立完成一条多肽链的合成,所以这种多聚核糖体可以在一条RNA链上同时合成多条相同的多肽链,这就大大提高了翻译的效率。
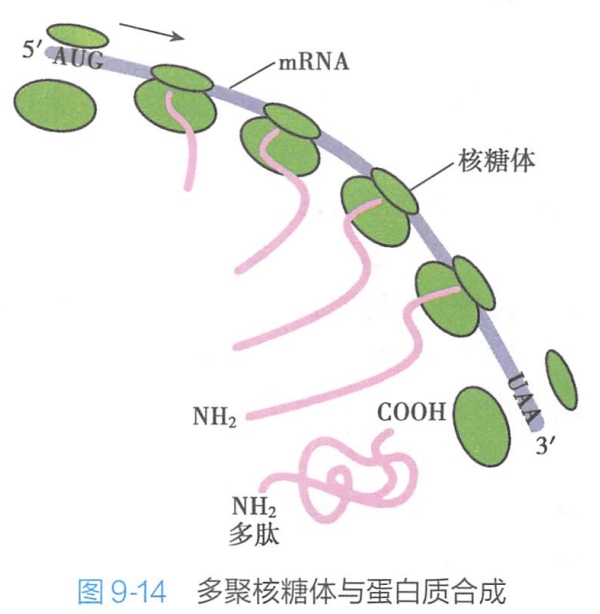
四、肽链合成后的加工修饰
新生多肽链尚不具有生物活性,必需经过化学修饰(如糖基化、甲基化等)和加工处理,使其在一级结构的基础上进一步盘曲、折叠,对于含有多个亚基的蛋白质还需要亚基与亚基间的结合。
(一)一级结构的修饰改变了多肽链氨基酸的性质和组成
新生多肽链一级结构的修饰常通过化学反应进行,如磷酸化、甲基化、二硫键形成等,这往往能改变多肽链氨基酸的性质和组成,并为空间结构的形成提供基础。
1.肽链氨基端的修饰
原核生物中几乎所有蛋白质合成都是从N-甲酰甲硫氨酸开始,真核生物从甲硫氨酸开始,但天然蛋白质大多数不是以甲硫氨酸或甲酰甲硫氨酸为氨基端起始的。因此,在多肽链合成终止,甚至在肽链合成进行的同时,甲酰基经脱甲酰基酶水解而除去,甲硫氨酸或者氨基端的一些氨基酸残基由氨肽酶水解掉,包括除去信号肽序列。
2.共价修饰
许多蛋白质可以进行不同类型化学基团的共价修饰,如磷酸化、糖基化、羟基化、甲基化、乙酰化和二硫键形成等,修饰后可以表现为激活状态,也可表现为失活状态。
- 磷酸化:磷酸化多发生在多肽链丝氨酸、苏氨酸和酪氨酸残基上,由蛋白激酶催化,磷酸化后蛋白质的活性发生改变;
- 糖基化:质膜蛋白质和许多分泌型蛋白质都具有糖链,这些寡糖链通过糖基化反应结合在丝氨酸或苏氨酸的羟基上,如红细胞膜上的ABO血型决定簇;
- 羟基化:胶原蛋白前体脯氨酸和赖氨酸残基羟基化形成羟脯氨酸和羟赖氨酸,对成熟胶原的链间共价交联是必需的;
- 二硫键:mRNA上没有胱氨酸的密码子,多肽链中的二硫键,是在肽链合成后,通过两个半胱氨酸的巯基氧化形成的,二硫键的形成对维持蛋白质的活性和结构是必需的。
3.多肽链的水解修饰
有些活性蛋白往往是由其无活性的前体蛋白水解而得来的,如大多数的蛋白酶原水解后转变为蛋白酶。
一般真核细胞中一个基因对应一个mRNA,一个mRNA对应一条多肽链,但少数情况下一种RNA翻译后的多肽链经水解后产生几种不同的蛋白质或多肽。
如哺乳动物的鸦片样促黑皮质激素原(proopio-melano-cortin,POMC)初翻译产物为265个氨基酸,
- 在垂体前叶细胞中,POMC初切割成为N-端片段和C-端片段的β-促脂解激素(β-LT,lipotropin)
- 然后N-端片段又被切割成较小的N-端片段和39肽的促肾上腺皮质激素(ACTH)
- 而在脑下垂体中叶细胞中,B-促脂解激素再次被切割产生β-内啡肽(β-endorphin);ACTH也被切割产生l3肽的促黑激素(α-melanotropin)
(二)高级结构的修饰
包括亚基聚合、多肽折叠和辅基连接。
1.亚基聚合
有许多蛋白质是由两个以上亚基构成,并需要各亚基肽链通过非共价键聚合成多聚体才能表现出完整的生物活性,如由两条α链、两条β链所组成的人血红蛋白、细胞膜上的镶嵌蛋白和穿膜蛋白等。
2.多肽折叠
新生肽链必须逐步折叠成天然空间构象才能成为有活性的功能蛋白。一般认为,蛋白质一级结构是其空间结构形成的基础,同时也需要其他的酶、蛋白质辅助才能完成折叠过程。分子伴侣(molecular chaperone)是一类能识别肽链的非天然结构、并能促进各功能域和整体蛋白质的正确折叠的酶或蛋白分子,它能介导其他蛋白质正确装配成有功能活性的空间结构,而本身并不参与最终装配产物的组成。目前认为“分子伴侣”蛋白有两类:
- 蛋白因子,如热休克蛋白(heat shock protein,HSP)、伴侣素(chaperonin),它们可以和部分折叠或没有折叠的蛋白质分子结合,稳定它们的构象,使其免遭其他酶的水解或促进蛋白质折叠成正确的空间结构;(内质网有网质蛋白)
- 酶,如蛋白质二硫键异构酶(protein disulfide isomerase,PDI),可以识别和水解非正确配对的二硫键;肽-脯氨酸顺反异构酶(peptide prolyl cis-.trans isomerase,PPI)能促进多肽链中肽酰-脯氨酸肽链的顺反异构体之间的转换。
3.辅基连接
有些蛋白,特别是结合蛋白和带辅基的酶,合成后都需要结合相应辅基,才能成为天然功能蛋白质。
五、蛋白质的降解
细胞内蛋白质在一系列蛋白酶的作用下水解成多肽,再在肽酶作用下降解成游离氨基酸,又再被重利用合成新的蛋白质。胞内蛋白质的降解主要通过两个途径,即泛素-蛋白酶体途径(ubiquitin proteasome system)和溶酶体途径。
(一)泛素-蛋白酶体途径
细胞中新合成的蛋白质中有超过30%需要通过泛素介导而被降解以保证蛋白质的质量。
泛素是1种含有76个氨基酸的小肽,被发现存在于大量不同的组织和有机体(但至今未在细菌中发现)中,因此命名为泛素(ubiquitin)。
泛素可以与待降解的蛋白质共价结合,而且一种蛋白质可以和多个泛素分子结合。后来将这种现象称为多泛素化(poly ubiquitination),蛋白质的多泛素化是启动蛋白质降解的信号。
在细胞内存在3种与泛素有关的重要的酶:泛素激活酶(E1)、泛素结合酶(E2)、泛素蛋白连接酶(E3)。
- E1激活泛素分子,此过程需要 ATP供能;
- 泛素分子被激活后被运送到E2上,E2负责将泛素绑在被降解的蛋白质上;
- E3能识别被降解的蛋白质;
- 最后,E3释放出被泛素标记的蛋白质。
- 如此循环反复,在结合了一定数量(一般认为至少5个)的泛素分子后,被降解蛋白质就被运送到细胞内的一种被称为“垃圾处理厂”的蛋白酶体(proteasome)结构中进行降解(图9-15)。
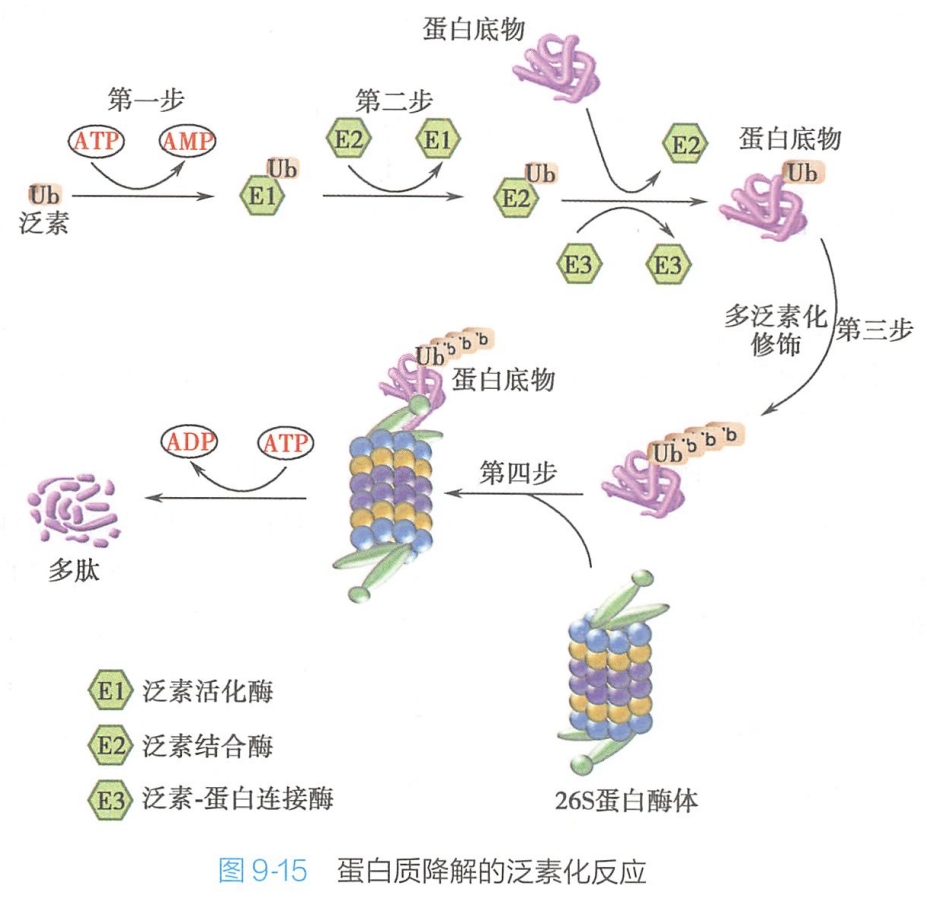
蛋白酶体:是一个26S蛋白复合体,主要存在于细胞核和细胞质中,负责依赖泛素的蛋白质降解途径,降解异常蛋白和短寿命蛋白。蛋白酶体本身不具备选择蛋白质的能力,只有被泛素分子标记而且被E3识别的蛋白质才能在蛋白酶体中进行降解。
(二)溶酶体途径
与蛋白质在蛋白酶体通过ATP依赖途径被降解的途径不同,溶酶体是真核细胞的消化器官,内含多种酸性水解酶,可以直接分解蛋白质分子。溶酶体降解蛋白质是一条非特异性的蛋白质降解系统,真核细胞内的膜蛋白、长寿蛋白和一部分短寿命蛋白都是在溶酶体中降解的。
蛋白质通过此途径降解,是不需要ATP提供能量的。
蛋白物质经自噬形成自噬体,或内吞作用进入细胞后,通过溶酶体消化,分解为小分子氨基酸。蛋白质在溶酶体的酸性环境中被相应的酶降解,然后通过溶酶体膜的载体蛋白运送至胞液,为合成新的蛋白质提供原料。